







ホーム > 修了生・同窓生の方 > 生物学科の学生活動報告(寄付金報告) > 奨学金受賞者(2025年度)
奨学金受賞者の研究紹介(2025年度)
2025年4月大学院博士後期課程に進学する優秀な学生に、この寄付金から奨学金が授与されました。受賞者の4人に研究内容をこのページで紹介してもらいました。植物発生研究室:長尾幸紀「アメリカネナシカズラの芽生えが葉及び根を形成しない分子機構の解明」
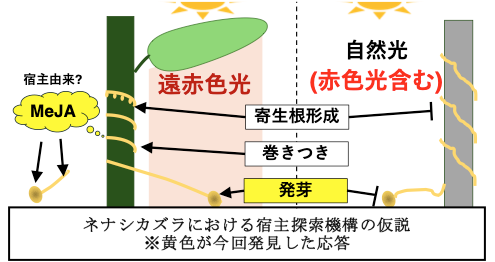
【研究の背景】植物の定義の一つに、「光合成を行う独立栄養生物」がありますが、寄生植物の中には光合成を行わないものも存在します。その一つである、ヒルガオ科に属するネナシカズラは根や葉の構造を持たず、茎だけの状態で寄生先の宿主に巻きつき、全ての栄養分や水分を彼らに依存する非常に興味深い生態を持っています。 宿主任せの生活であるが故に、ネナシカズラは寄生に失敗すると発芽後10日程度で枯れてしまいます。しかし周囲に植物が茂った環境では、寄生根と呼ばれる器官を形成して宿主に侵入し、自身と相手の維管束同士を結合させることで養分を吸い取り、急速に成長します。中でもアメリカネナシカズラ(Cuscuta campestris)は外来種として分布域を拡大し、農作物や海浜の在来植物へ被害を与えることが報告されており、駆除方法の検討が喫緊の課題です。

【卒業/修了研究の内容】
アメリカネナシカズラの生態を理解するために、種子発芽と芽生えの成長について生理学的解析を行いました。その結果、発芽及び成長に対して一般的な植物では抑制的に作用するジャスモン酸に促進効果があることや、遠赤色光が発芽を促進し、フィトクロムを介した赤色-遠赤色可逆応答が光発芽種子や暗発芽種子と異なることを発見しました。寄生根の形成や巻きつき運動に、遠赤色光が重要である例は報告されています。それに加え、発芽の段階から、宿主由来のジャスモン酸や宿主の葉が太陽光から吸収する波長を除いた光、すなわち遠赤色光を用いて宿主を探索するシグナルとして利用している可能性があります。また、サイトカイニンの添加処理により、通常は寄生後に誘導される分枝構造や鱗片葉の形成、オーキシン添加処理により、発芽後すぐに退化、枯死する根及び根端の毛状細胞を維持することに成功しました。
【今後の研究予定】
得られたホルモン応答の知見から、ネナシカズラがなぜ根や葉を失い「根無蔓」になったのかを遺伝学的手法で明らかにしていきたいと考えています。また、未だ確立されていないネナシカズラにおけるゲノム編集技術の開発にも並列して取り組む予定です。
分子内分泌学研究室:竹内優羽「光周性の制御リレーを担う脳内の組織構造の解析」
下垂体隆起部は,下垂体前葉が口吻側に薄く伸びた領域で,脳底側から正中隆起を覆うように存在します(図1)。哺乳類では隆起部にメラトニン受容体が高発現するため,1日の光周期(日照時間)の情報は,メラトニンを介して隆起部ホルモンの分泌リズムに変換され,生理機能に日周的・季節的なリズムが生まれていると考えられてきました。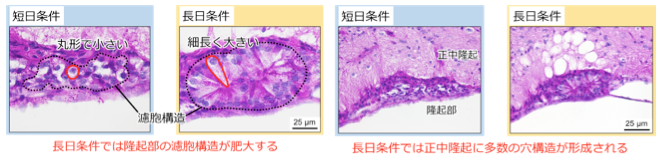
図1 下垂体隆起部
実際に,繁殖に季節性を示すヒツジやハムスターでは,隆起部の甲状腺刺激ホルモン(TSH)が日照時間の長い長日条件で劇的に増加し,第三脳室の特殊な上衣細胞(タニサイト)の形態変化を引き起こすことで,性腺刺激ホルモン放出ホルモン(GnRH)の分泌制御を担うことが知られています。 しかしながら,1年中繁殖が可能なラットではこのような季節性に着目した研究は少なく,未解明な部分も多く残されています。そこで私は,実験動物ラットにおいて光周期の変化が隆起部を含む脳の組織構造にどのような影響を及ぼすのかを明らかにすることを目的として研究を進めています。
F344系統のラットを長日条件下で飼育すると,驚くべきことに隆起部内の濾胞構造を形成する細胞が通常の2倍にまで肥大すること,隆起部近傍に位置する正中隆起に多数の穴構造が出現することが明らかになりました(図2)。長日条件下でラットにメラトニンを投与した場合には,これら脳の組織構造の変化は起きなかったことから,短日条件および通常の光周期ではメラトニンによって抑制されていると考えられます。
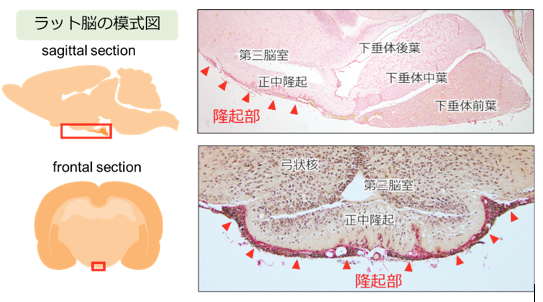
図2 長日条件下で観察される脳の組織構造の変化
正中隆起は視床下部の向下垂体ホルモン産生神経の軸索が通る重要な領域です。その一つである性腺刺激ホルモン放出ホルモン(GnRH)ニューロンの軸索の一部が穴構造の周囲には密集する様子が観察されました。下垂体前葉において性腺刺激ホルモン(LH)のLhb mRNAの発現が減少していることから,長日条件で形成される正中隆起の穴構造がGnRHの分泌を抑制する機能があることが示唆されました。
博士後期課程では正中隆起の穴構造の形成メカニズム,そして生理的意義を解明すべく研究を進めていきます。正中隆起を通る向下垂体ホルモン産生神経ではGnRHニューロンのほかに,チロシン水酸化酵素(TH)ニューロンの軸索も穴構造の周囲に密に存在しました。THはドーパミン産生の律速酵素であり,ドーパミンは下垂体前葉に作用しプロラクチン(PRL)の分泌を抑制します。つまり,穴構造はTHニューロンになんらかの効果をもたらし,PRLの分泌を制御する組織構造である可能性があります。今後は穴構造とPRLとの関係性にも着目したいと考えています。
生体超分子構造研究室:Jiang Haowei「光合成タンパク質Ⅱ変異体において水分子分解と活性中心変化解明」
【研究の背景】生命の進化の過程において、光合成は非常に重要な役割を果たしてきました。光合成の中に重要な機能を担うのは、Photosystem II(PSII)と呼ばれるタンパク質です。多数の色素分子と複雑な構造を有しており、これらの色素分子は太陽光エネルギーを吸収し、それを段階的に反応中心へ伝達します。そして反応中心は得られたエネルギーを用いて水分子を分解し、酸素を発生するとともに、同時にプロトンと電子を生成します。私はPSIIの回転サイクルおよび活性中心の解明を目的として、研究しています。
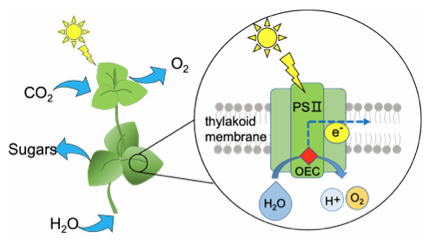
図1.PSIIの水分解
【研究内容】
私はクライオ電子顕微鏡を用いて、野生型および暗条件下での変異体タンパク質を観察しながら、PSIIのメカニズムを解析しています。具体的には、PSIIの活性部位近傍に存在する185番目のバリン(Val)をスレオニン(Thr)に置換した変異体を作製しました。この変異体は野生型のわずか13%程度の活性しか示さないにもかかわらず、外部エネルギーを利用しなくても光合成のみで成長できることが明らかになっています。そして、観察した変異では本来安定していたアミノ酸残基が不安定化し、変異体PSII中では変異点が2種類のコンフォメーションとして観察されます。そのうちの1つのコンフォメーションは、PSIIが周辺部から活性部位へ水を輸送する通路に影響を与え、もう1つのコンフォメーションは水の分解後にプロトンを排出するチャネルを断裂させる可能性が示唆されています。これらの結果は、このアミノ酸残基が活性部位に疎水的環境を与え、それを安定化するうえで重要な役割を担っていることを意味しています。さらに、構造解析からはV185が複数のチャネルを安定化し、Mnクラスターに対して疎水性を付与することで正常な機能を支えていることも明らかになりました。
【今後の展望】
この変異体はS2 High-spin Stateと測定しましたが、構造的には解明されていないです。そのHigh-spin状態では光1回Flash後に観察できることを期待しています。それで、私はPSIIに1回を光照射して、High spin状態を観察し、構造を解明できるように頑張ります。
分子機能研究室:大常真一「オスのショウジョウバエにおける内部生殖器の発生制御機構の解明」
【研究の背景】少子化が進む現代において,不妊症は深刻な課題の一つです.男性の生殖機能に着目すると,妊娠能力には精子の数は十分であるか,精液成分に含まれるタンパク質は正常に分泌されているかが大きく影響します.
【研究内容】
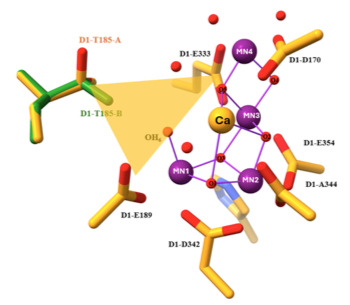
ショウジョウバエ附属腺は,ヒトの精嚢や前立腺と相同な内部生殖器官で,約1000個の主細胞と約60個の第二細胞 (SC) から構成されており,それぞれが精液成分を分泌しています.成虫のSCで,ホメオドメイン型転写制御因子 Dve が発現しています (図1).dve 変異体では SCが消失し,オスの妊娠能力は大幅に低下します.さらに,栄養環境に応じて妊娠能力が変動しますが,SCの大きさも連動して変化します.つまり,附属腺において主細胞のわずか6%に過ぎないSCが,適切な妊娠能力に必須であり,外部環境に応答した適応的な生殖を可能にしていることになります.
図1. ショウジョウバエの附属腺
附属腺の正常な発生には,ステロイドホルモンであるエクダイソンが必要であるため,エクダイソンシグナルと SCの大きさ制御の関連性について解析を行いました.表皮においてエクダイソンシグナルの下流因子として知られるpriの発現を附属腺で阻害すると,SCが小さくなることがわかりました.さらに,priの強制発現によって,エクダイソンシグナルの阻害によるSCの萎縮が改善されることもわかりました.続いて,すでにエクダイソンシグナルの下流因子として知られているFtz-F1の下流で pri が機能することを明らかにしました (図2).
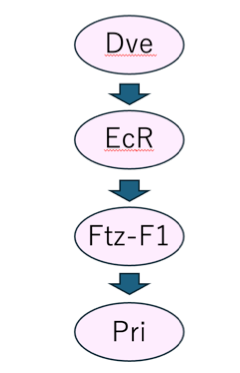
図2. SCの大きさ制御機構
【今後の研究】
博士課程後期では,主細胞が担う機能 (精子の運動性など) にも着目し,エクダイソンシグナルがオスの生殖能力をどのように制御するのかを解明していきます.得られた知見を哺乳類に活かすことができたなら,とても嬉しく思います.
